人类神经系统发育详尽报告——谷歌gemini2.5 deepsearch
人类神经系统发育详尽报告
注: 《有关神经系统科学基本原理的信息》,谷歌gemini2.5这个研究报告有详细的分子过程。下一步我们将转向有关“连接组学”的研究。
人类神经系统是生物学中最复杂的结构之一,其发育过程是一个从受精后不久开始,持续到成年期的漫长而精密的旅程。这一过程涉及神经元和胶质细胞的精确生成、迁移、分化、成熟、突触形成与修剪以及髓鞘化,所有这些都由复杂的内在遗传程序和外在环境线索所引导。对这一复杂发育过程的深入理解,对于揭示脑功能、诊断和治疗神经发育障碍至关重要。
一、引言
人类神经系统发育概述
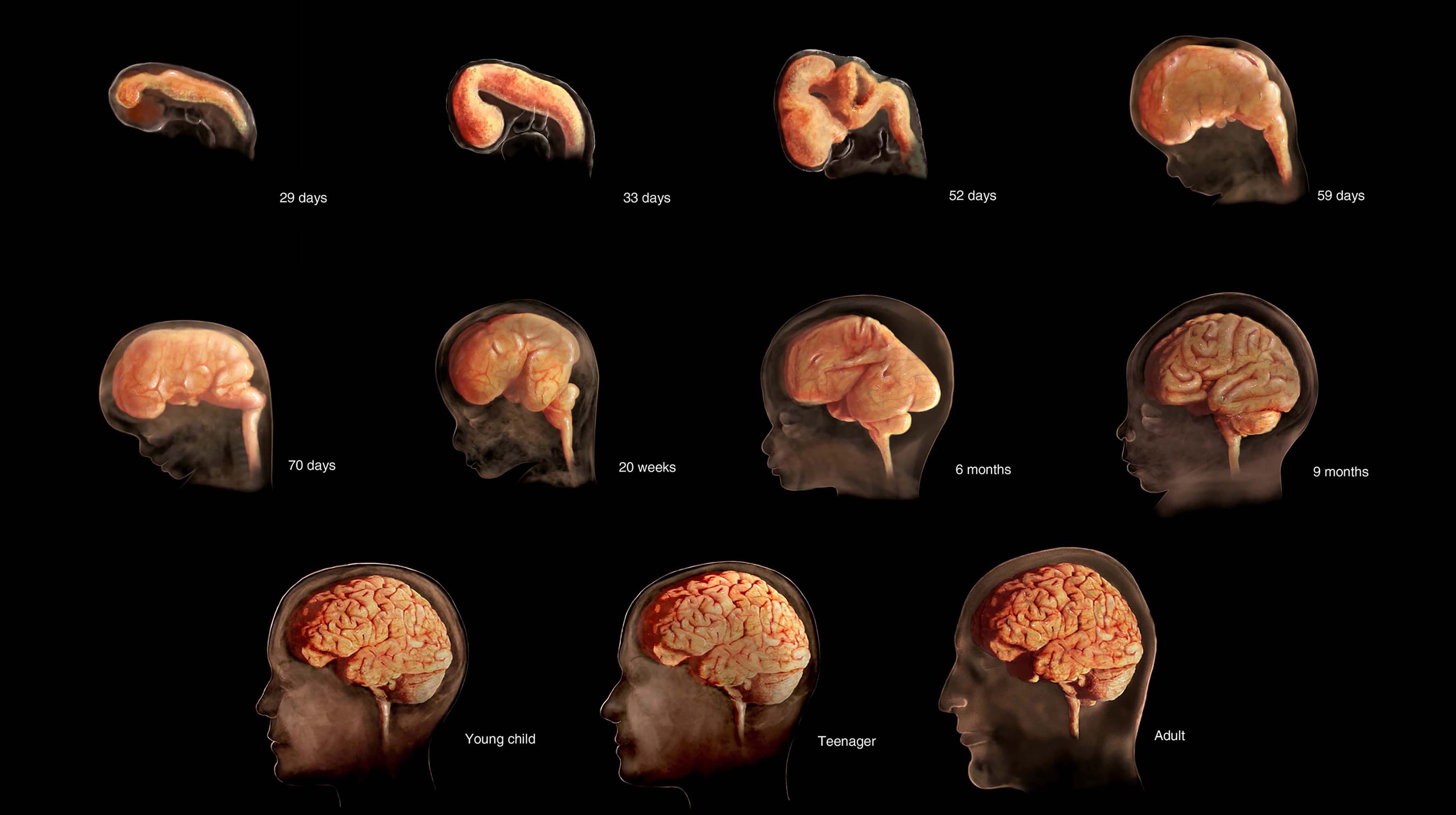
人类神经系统从一个简单的外胚层细胞层开始,逐步演变为高度组织化的大脑和脊髓,这一过程是生物学复杂性的典范 1。神经系统的发育是一个漫长且精确协调的旅程,它在受孕后不久启动,并持续到成年期 1。这个错综复杂的过程包含了一系列基础性的细胞事件,包括神经发生(神经元生成)、细胞迁移、细胞分化、细胞成熟、突触发生(突触形成)、细胞死亡与修剪,以及髓鞘形成 4。
意义与总体时间线
神经系统的原基最早在受精后第三周出现,但其分化和成熟需要相当长的时间,直至出生后很长一段时间 2。关键的里程碑包括:神经管在大约受孕后第四周闭合;神经元快速增殖(神经发生),在受孕后约100天达到峰值,每小时生成约1500万个神经元 6;出生时,大脑大小约为成人大脑的四分之一 8。尽管大脑在约14岁时达到其物理上的完全大小,但关键的神经回路重塑,包括广泛的髓鞘形成过程,会持续到成年早期,通常在25至30岁左右才完全成熟 3。
神经系统发育的这种漫长轨迹,意味着其在结构、连接和功能能力方面,对遗传倾向、环境因素(如营养、毒素、压力)和经验输入(如社会互动、学习)等多种影响因素,具有持续且延长的敏感性窗口 6。这意味着,即使在婴儿期之后发生的损伤或缺陷,仍可能对神经系统的最终形成和功能产生深远影响,从而塑造个体一生中的认知能力、情绪调节和整体健康状况。因此,持续的支持性环境、全面的营养支持以及避免不利因素的暴露,不仅在儿童早期,而且在整个青少年期和成年早期都至关重要,以促进最佳的神经功能。
表1:人类神经系统发育的关键阶段及其时间线
| 阶段 | 大致时间线(受孕后周数/出生后年龄) | 关键事件/结构 |
|---|---|---|
| 神经诱导与神经管形成 | 2-4 WPC | 神经板、神经沟、神经管形成;神经孔闭合 |
| 原始脑泡形成 | 3-4 WPC | 前脑、中脑、菱脑分化 |
| 神经发生(高峰期) | 4 WPC - 18个月 PNA | 神经元大量生成(约460万个/小时);脑室区和室管膜下区 |
| 神经元迁移 | 12-20 WPC | 新生神经元从出生地向最终目的地移动;放射状和切向迁移 |
| 突触发生(起始/旺盛期) | 第三孕期 - 2岁 PNA | 突触形成爆发性增长;皮质深层和脑干区域先于浅层 |
| 胶质发生(起始) | 24 WPC - 儿童早期 | 星形胶质细胞、少突胶质细胞、小胶质细胞出现 |
| 髓鞘化(起始/完成) | 第一孕期末(脊髓)- 25-30岁 PNA | 髓鞘包裹神经纤维;脊髓、脑干先于大脑皮层 |
| 突触修剪(高峰期) | 儿童早期 - 青少年期 | 消除不必要或功能不良的神经元和突触 |
二、神经系统的早期胚胎基础
A. 神经诱导与神经管形成:从外胚层到神经管
中枢神经系统(CNS),包括大脑和脊髓,起源于胚胎最外层的胚层——外胚层 1。大约在受精后第二周,背侧外胚层的特定区域被指定为神经外胚层,随后增厚形成神经板 1。这是胚胎早期模式形成的关键步骤,建立了前后轴 1。
在大约受孕后第18天,神经板沿其中轴内陷,形成神经沟,两侧是双侧神经褶 5。这一折叠过程被称为神经管形成(neurulation),最终神经褶融合形成神经管 1。这种融合通常始于未来颈部区域,并向颅侧(头侧)和尾侧延伸 5。神经管的前(头端)和后(尾端)神经孔分别在大约第24天和第28天闭合,完成神经管的形成 5。神经管的空心内部随后充满胚胎脑脊液 1。
神经板诱导的分子机制
未分化外胚层向神经外胚层的转化,关键依赖于下方中胚层的诱导信号 1。在原肠胚形成开始时,推定的中胚层细胞通过背侧胚孔唇移动,在中内胚层之间形成一层中胚层 1。脊索,作为一种关键的中胚层结构,通过分泌拮抗分子如
卵泡抑素(Follistatin)、合体素(Chordin)和诺金(Noggin),在神经诱导中发挥核心作用 11。这些分泌因子特异性地抑制骨形态发生蛋白(BMPs),特别是BMP4的活性,否则BMP4会抑制外胚层细胞向神经细胞的分化 11。最近的研究还表明,成纤维细胞生长因子(FGFs)在早期阶段就能抑制BMP4的表达 11。脊索还诱导神经管的腹侧部分,即基板,其将形成运动结构 1。相反,神经管的背侧区域,即未来的感觉顶板(翼板),则由TGF-β家族(如Dorsalin、Activin、BMP4)和Wnts等一系列生长因子进行背侧化 11。
神经诱导过程的根本机制并非通过开启一个全新的通路来直接“诱导”神经命运,而更像是一种“解除抑制”的机制。外胚层的默认发育路径是神经元,但这种命运被BMP信号传导积极地抑制。中胚层的作用是分泌BMP拮抗剂,从而解除这种“制动”,允许内在的神经程序得以进行 11。例如,诺金通过与BMPs紧密结合,阻止BMPs与其受体结合 12。这种对默认通路的深刻理解是发育生物学的基石,对理解发育错误如何发生具有重要意义。BMPs与其拮抗剂之间微妙平衡的破坏,或脊索诱导信号的异常,即使外胚层细胞本身具有成为神经元的内在能力,也可能导致严重的神经模式形成缺陷(例如,神经板形成失败或神经管背腹侧模式形成不当)。这一机制强调了早期胚胎发生中精确分子反向调节的重要性。
B. 原始脑泡和次级脑泡的形成
随着神经管的闭合,其前(头侧)部分显著扩张,形成三个原始脑泡:前脑(prosencephalon)、中脑(mesencephalon)和菱脑(rhombencephalon) 1。这些基本分区在进化上是保守的,并存在于所有脊椎动物中 13。
这些原始脑泡随后进一步分裂和分化,形成更专业的脑区。前脑分为端脑(telencephalon)(将形成未来的大脑皮层、海马和基底神经节)和间脑(diencephalon)(将形成未来的丘脑和下丘脑) 1。值得注意的是,最终发育成视神经、视网膜和虹膜的视泡,形成于前脑的基板 1。菱脑进一步分为头侧部分——
后脑(metencephalon)(形成未来的脑桥和小脑),和尾侧部分——脊髓脑(myelencephalon)(形成未来的延髓) 1。中脑通常保持为单一的未分化结构 1。
发育中的神经管也表现出特征性的弯曲,包括中脑弯曲(在中脑水平)、颈曲(在菱脑和脊髓的连接处)以及脑桥弯曲(在后脑内) 1。这些弯曲对发育中的大脑的整体复杂形状和组织起着重要作用。神经管最初的空心内部持续存在并扩大,形成包含脑脊液的脑室系统 1。
三、神经发育的基本细胞过程
A. 神经发生与神经祖细胞动力学
神经发生,即生成新神经元的过程,是一个基础性的事件,主要发生在胚胎发育期间,尽管在某些特定的成人脑区仍有少量持续发生 4。这一过程始于胚胎发育的第四周左右,神经干细胞在神经管的脑室区(VZ)内增殖 2。这些神经干细胞是特殊类型的多能细胞,主要生成放射状胶质细胞(RGCs),而RGCs又会产生中枢神经系统中的各种神经元和胶质细胞 2。人类端脑(包括皮层、海马和基底神经节)的绝大多数神经元在出生前生成 14。然而,在其他脑区,特别是小脑,出生后也发生广泛的神经发生 7。在这一早期发育阶段,即受孕后第四周至出生后18个月之间,中枢神经系统以每小时约460万个神经元的惊人速度生成神经元 14。
最初,排列在脑室内的神经上皮细胞(NECs)通过对称分裂来扩大总的祖细胞池,这种机制显著影响新皮层的最终表面积和厚度 2。在神经发生开始时,NECs转变为顶端放射状胶质细胞(aRGCs)。这些aRGCs保留了神经上皮特征,包括黏附连接和顶基极性,并延伸出长长的基底突起,跨越整个皮层厚度,作为神经元迁移的支架 2。aRGCs可以通过对称分裂进行自我更新,但主要通过不对称神经发生分裂,产生一个新的aRGC和直接的神经元(直接神经发生)或一个中间祖细胞(IPC)(间接神经发生) 2。IPCs是瞬时扩增的祖细胞,其特征是多极形态并表达转录因子Tbr2 2。它们从脑室区脱离并定居在室管膜下区(SVZ),在那里进行终末对称分裂,产生两个神经元 2。因此,SVZ成为主要的生发基质,产生大多数皮层细胞 7。
细胞分裂和命运的分子调控
神经发生与随后的胶质发生之间的精确时间和平衡,通过内在因素(例如,基因表观遗传修饰)和外在线索(分泌或接触介导的因子)之间复杂的相互作用进行精细调控 2。
核间运动(INM): NECs和脑室区祖细胞的一个独特特征是核间运动(INM),其中细胞核在细胞内的物理位置与细胞周期阶段协调一致。细胞核在S期向基底端迁移,在G2期向顶端迁移,在脑室表面进行M期,并在G1期返回基底端 2。这种高度协调的运动,由与微管相关的运动蛋白复合物(例如,动力蛋白、Lis1、Nde1用于基底到顶端迁移;肌球蛋白II用于顶端到基底迁移)驱动,对于维持祖细胞池的稳态至关重要 2。INM的损伤可能导致异常的脑室外有丝分裂、细胞凋亡或过早退出细胞周期,从而导致显著的祖细胞损失 2。Notch信号通路: Notch信号通路在祖细胞命运调控中发挥关键作用。Notch信号通路通过高表达的Par3维持,促进祖细胞增殖并抑制神经发生基因(例如,Hes基因),从而抑制过早的分化分裂 2。在不对称分裂过程中,Par3的不均匀分配导致继承较少Par3的子细胞退出细胞周期并分化 2。Notch信号通路还受到脉络丛分泌到脑脊液中的因子(包括Igf2、FGFs、Wnts和Shh)的进一步调节 2。糖原合成酶激酶3(GSK3): GSK3已知会抑制神经祖细胞增殖,其失活通过各种信号通路的相互作用得以维持 2。基因调控与畸形: 许多基因控制着神经发生,调节转录、细胞周期进程(例如,MCPH1、CENPJ、CDK5RAP2)、中心体成熟、DNA修复(例如,PNKP、PCNT)以及神经干细胞的增殖能力(例如,ASPM和STIL) 7。这些关键基因的突变可能导致严重的先天性小头畸形,其特征是由于神经元和胶质细胞增殖减少而导致脑体积减小 2。相反,神经祖细胞的过度增殖可能导致巨头畸形,这突显了正常脑体积所需的精确控制 2。
人类大脑中大量神经元和多样性的产生,关键在于生发区内细胞分裂的高度精确的时空控制。细胞分裂的物理力学,包括细胞核定位(INM)和有丝分裂纺锤体的方向(对称与不对称分裂),是根本的调控机制。这些物理过程与Notch等分子信号通路协同作用,确保了祖细胞的正确数量、适当扩增以及及时分化。对这种物理组织的破坏可能对大脑发育产生深远而直接的影响,导致小头畸形或巨头畸形等疾病,即使细胞类型的遗传指令在其他方面是完整的。这一认识强调了神经发育不仅是一个生化级联反应,也是一个生物物理过程。发育中的大脑内的细胞结构和机械力在塑造其最终形态和功能方面发挥着积极和指导作用。这种理解可能为某些神经发育障碍的物理基础研究开辟新途径,并可能为针对细胞力学或组织组织而非仅仅分子通路的治疗策略提供信息。
B. 神经元迁移:引导神经元到达目的地
神经元在神经发生后,从其增殖的出生地(主要是脑室区和室管膜下区)开始精确而广泛的旅程,到达发育中大脑中其最终的指定位置 1。这种定向运动,被称为神经元迁移,对于脑回路的适当布线和不同脑结构的自组织至关重要 1。
放射状和切向迁移路径
神经元迁移主要通过两种模式进行:
放射状迁移: 在这种模式下,神经元垂直于神经上皮表面移动,通常利用放射状胶质细胞(RGCs)的长突起作为物理支架或引导 7。这是大脑皮层径向延伸和层状组织的主要过程,其中较年轻、晚期出生的神经元会迁移过较老、早期出生的神经元,占据更表层的区域,从而建立皮层发育的“由内向外”模式 2。切向迁移: 相比之下,切向迁移涉及神经元平行于软脑膜表面(大脑表面)移动,由各种可扩散和膜结合的分子线索引导 10。这种模式对于来自不同增殖区(通常具有不同谱系)的神经元汇聚并形成远端脑区中的适当突触连接至关重要 15。
关键引导线索和受体
迁移神经元的精确导航由复杂的化学信号和分子相互作用阵列所协调:
Reelin: 由发育中皮层边缘带(第一层)的Cajal-Retzius细胞分泌,Reelin是一种关键的细胞外基质糖蛋白 10。它在皮层分层中发挥关键作用,并被认为是放射状迁移神经元的“脱离和停止信号”,帮助它们在到达最终目的地时脱离放射状胶质细胞 10。基因研究表明,Reelin通路中的突变,包括其受体(ApoER2、Vldr)和衔接蛋白(Dab1),会导致皮层分层的严重破坏 10。Reelin还诱导迁移模式从运动模式转变为终末体易位模式,当神经元接近其目标层时 16。信号素(Semaphorins)、神经纤毛蛋白(Neuropilins)、细胞黏附分子(Cell Adhesion Molecules)、神经调节蛋白(Neuregulins)、趋化因子(Chemokines)、Slit和Robo家族: 这些多样化的分子家族是主要参与指导切向迁移的关键引导线索 10。例如,Slit及其受体Robo对于阻止轴突在最初响应其他向性信号穿越中线后再次穿越中线特别重要 17。信号素可以作为排斥性和吸引性引导分子,具体取决于激活的特定受体复合物、神经元亚型和发育时间 18。细胞外基质成分: 除了直接的细胞间相互作用,细胞外基质的成分也作为神经元运动的底物,特别是在体易位期间 19。作为引导线索的神经递质: 有趣的是,神经递质如γ-氨基丁酸(GABA)和谷氨酸,由神经元自身分泌,可以作为外在因子,加速神经元迁移或刺激皮层中间神经元的运动性 19。细胞内钙(Ca2+)内流: 细胞内钙的增加是细胞到达最终目的地后停止神经元迁移的提议机制,标志着其迁移程序的结束 10。
迁移周期本身涉及神经元前导过程的动态变化和细胞体(体易位)的运动,这关键依赖于微管阵列和中心体的完整性和动力学 10。一旦神经元到达其区域位置,它们必须停止其迁移程序并分化成其专门形式 1。神经元迁移过程中的异常是各种中枢神经系统畸形的重要潜在原因 7。
神经元迁移和轴突寻径的引导并非由简单的单向“开/关”或“吸引/排斥”信号控制。相反,它涉及高度复杂、通常是双向且依赖于情境的信号机制。单个分子线索(例如,Reelin、信号素)可以根据其浓度梯度、迁移神经元上的特定受体环境、共存因子的存在、发育阶段或特定的神经元亚型,发挥多种、有时看似矛盾的作用 10。这种错综复杂的相互作用使得大脑中观察到的布线模式具有卓越的精确性和多样性。对这种情境依赖性的深刻理解对于基础神经科学和临床应用都至关重要。例如,在理解迁移受损的神经发育障碍时,如果未能充分认识到单个引导分子的多方面作用及其与其他信号的复杂相互作用,简单地增加或减少其表达可能无法达到预期的治疗效果。这强调了在尝试调节发育过程时,考虑整个分子和细胞微环境的挑战性和必要性。
C. 神经元分化与成熟
在成功迁移之后,神经元进入分化阶段,在此阶段它们变得专门化并获得独特的功能 4。在这个关键时期,神经元开始表达特定的基因,这些基因决定了它们的独特身份以及它们在大脑回路中的精确作用 4。作为成熟过程的一部分,神经元经历显著的结构和功能变化,包括形态上的巨大改变,例如延伸出复杂的轴突和树突过程 1。这些延伸对于建立神经元之间错综复杂的连接网络至关重要,而这个网络是神经通信的基础 1。
D. 突触发生:建立神经连接
突触发生,即突触的形成——允许神经元之间进行通信和信息处理的特化连接——是神经系统发育中一个基本且精确计时的过程 4。它代表了轴突寻径的最终也是关键阶段,其中轴突尖端或生长锥接近其目标神经元以形成功能性连接 20。
突触形成的时间线和模式
大脑的生长和发育涉及突触形成的“爆发”,称为旺盛突触发生,这在早期大脑发育中显著发生 21。这一过程始于妊娠第三期,并广泛持续到出生后的头两年 21。
在人类大脑皮层中,在约20周妊娠期(GA)时,在边缘带(分子层)的Cajal-Retzius神经元横向轴突中仅观察到极少的突触素反应性(突触囊泡的结构糖蛋白标记物) 20。到26周GA时,分子层和较深的皮层层显示出显著的突触素反应性,而浅层2则基本无反应 20。在33周GA时,皮层深层表现出比浅层更强的反应性,表明突触成熟的进展 20。在小脑中,关键结构(包括齿状核、红核和下橄榄核组成的Guillain-Mollaret三角)的突触发生在胎儿发育早期就显著发生 23。最早在13周GA时,在红核和下橄榄核中检测到初始突触囊泡,在16周GA时在齿状核中检测到 23。这突出强调了发育神经解剖学的一个基本原则:突触形成可能先于这些核团达到成熟形态 23。关于特定的神经递质系统,谷氨酸能(兴奋性)神经元在大约9周GA时开始在锥体细胞中产生突触活动 24。已知抑制性神经元在发育中的皮层内进行切向迁移 24。皮层下板在28周GA时高度活跃,启动了显著的突触发生和早期突触可塑性,主要发生在额叶和顶叶区域 24。多巴胺能神经元,对运动、动机和奖励行为至关重要,在大约6-8周GA时开始其迁移和在突触发生中的作用 24。
突触组装的分子成分
突触发生依赖于众多分子成分的精确组装。突触素是一种主要的结构糖蛋白,是突触囊泡膜的主要组成部分,也是证明突触发生序列的可靠标记物 20。这些囊泡膜的其他关键结构糖蛋白包括突触结合蛋白(synaptobrevin)和突触结合蛋白(synaptotagmin),其中后者对于微摩尔级突触前钙信号介导的突触囊泡快速补充尤为重要 20。此外,EphB与Ephrin-B的结合导致双向信号传导,这在发育过程中参与轴突引导和边界形成,并已被证明可以调节突触后组织 21。
突触形成的时间或分布异常,包括兴奋性和抑制性影响比例的改变,可能导致皮层发育畸形和皮层畸形中癫痫发生率增加等病理结果 20。
尽管大脑在出生后会经历突触连接的大规模扩张和精细化,但基础的突触回路和最初的功能活动在胎儿生命早期就已经建立 20。例如,谷氨酸能神经元在妊娠第9周左右就表现出突触活动,而皮层下板在第28周左右就显示出高度的突触发生和早期突触可塑性。这表明,功能连接的形成,尽管是初步的,但早于胎儿后期和出生后早期广泛的“旺盛突触发生”和修剪阶段。这一观察强调了早期功能特化在神经发育中的重要性。这些早期形成的连接为后续更复杂的回路发展奠定了基础,并可能决定了大脑处理信息和应对环境刺激的初始能力。因此,任何在这些早期阶段发生的干扰都可能对后续的神经回路成熟和功能产生级联效应,从而影响个体的认知、情感和社会发展。
四、胶质细胞发育与髓鞘化
A. 胶质发生:支持细胞的诞生
胶质发生是指胶质细胞的生成过程,其时间晚于神经发生,从胚胎晚期持续到儿童早期 2。星形胶质细胞和少突胶质细胞起源于神经上皮,而小胶质细胞则起源于卵黄囊中的造血干细胞 25。从神经发生向胶质发生的转换涉及促神经因子下调和促胶质转录程序的激活 2。
B. 少突胶质细胞发育与髓鞘化
少突胶质细胞的作用
少突胶质细胞(OLGs)是中枢神经系统(CNS)中形成髓鞘的特化胶质细胞,它们通过包裹神经轴突形成髓鞘,从而显著提高电信号的传导速度和效率 3。髓鞘作为一种脂质丰富的绝缘膜,通过减少离子泄漏和降低细胞膜电容,使动作电位沿轴突的跳跃式传导成为可能,从而实现比相同直径的无髓鞘轴突快20-100倍的神经传导速度 3。除了提高传导速度,髓鞘还通过乳酸转运为轴突提供代谢支持 30。
少突胶质前体细胞(OPC)发育
少突胶质细胞源自少突胶质前体细胞(OPCs),OPCs主要在胚胎期腹侧脑室区域(如内侧神经节隆起,MGE)生成,随后在侧神经节隆起(LGE)和皮层中生成 2。OPCs增殖并迁移到大脑皮层,在纤维束中分化为髓鞘前少突胶质细胞(preOLs),最终成熟为髓鞘化少突胶质细胞 2。
髓鞘化时间线
髓鞘化是一个漫长而分阶段的过程。在人类中,髓鞘化最早在妊娠第一孕期末开始于脊髓 5。在胚胎第8周,脑干开始髓鞘化 5。随后,髓鞘化从妊娠32周持续到成年期,在出生后6个月到2岁达到高峰 2。整个过程通常要到25-30岁才完全完成 2。髓鞘化进展呈现出从背侧到腹侧、从尾侧到头侧的梯度 2。髓鞘的形成与儿童的智商测试结果呈正相关,表明其对认知功能的重要性 3。
少突胶质细胞特化/分化的分子调控
少突胶质细胞的特化、增殖、存活、迁移、分化和髓鞘化受到多种分子机制的精确调控:
信号分子: Sonic Hedgehog (Shh) 对早期腹侧前脑OPC发育至关重要 2。BMP和Wnt/β-catenin通路则拮抗Shh,并负向调节少突胶质细胞的诱导,其中BMP4促进星形胶质细胞谱系,Wnt/β-catenin早期抑制OPC生成,后期促进少突胶质细胞分化 2。Notch信号通路抑制神经元和少突胶质细胞分化,同时促进星形胶质细胞分化 2。促有丝分裂信号: 血小板衍生生长因子-A(PDGF-A)、FGF家族成员、神经营养因子-3、胰岛素和胰岛素样生长因子(IGFs)促进OPC增殖和存活,但抑制其分化 2。细胞内在因子和转录因子: 细胞内在因子控制OPC从增殖向分化的转变。甲状腺激素T3可能作为外在共信号。转录因子如Olig1/2、Sox10、Nkx2.2、Myrf、Sip1、Mash1是正向调节因子 2。而LINGO1、GPR17、Hes5、Id2/Id4则是负向调节因子 2。表观遗传因子: miRNA(miR-219、miR-338)、非编码RNA和染色质重塑分子(HDACs、Dnmt3a、Amarca4/Brg1)调节少突胶质细胞的分化 2。
髓鞘化的分子调节因子
Fyn激酶: Fyn激酶在CNS髓鞘化中发挥独特作用,调节少突胶质细胞的成熟、突起生长和髓鞘碱性蛋白(MBP)mRNA的转运 2。轴突影响: 轴突接触、直径、黏附分子(PSA-NCAM)和神经营养因子调节髓鞘化 2。神经元活动: 兴奋性神经元活动通过与OPCs/preOLs的突触以及谷氨酸/ATP的释放,作用于AMPA或嘌呤能受体,从而影响少突胶质细胞的发育 2。轴突-OPC突触在发育中的大脑白质损伤中高度脆弱,早产儿的preOLs瞬时表达缺乏GluR2的Ca2+通透性AMPA受体,使其在缺氧-缺血条件下易受兴奋性毒性影响 2。preOLs也对氧化应激和炎症敏感,其死亡和成熟失败导致早产儿大脑髓鞘不足 2。
髓鞘化是一个动态的、依赖于神经元活动的复杂过程,受到神经元信号(如谷氨酸、ATP、神经营养蛋白等生长因子)和胶质细胞间相互作用的深刻影响 9。这挑战了髓鞘仅作为静态绝缘体的传统观念,揭示了其在神经可塑性和高级认知功能中的积极作用。例如,神经元活动可以促进髓鞘化,表明髓鞘的形成和重塑是适应性过程,能够响应神经回路的需求。这种动态特性对于理解大脑如何学习和适应至关重要,并为治疗髓鞘疾病提供了新的靶点,可能通过调节神经元活动或神经元-胶质细胞信号传导来促进髓鞘修复。
C. 星形胶质细胞发育与功能
起源与时间
星形胶质细胞(Astrocytes)是中枢神经系统中最丰富的细胞类型 25。星形胶质细胞的生成(astrogenesis)发生在神经发生之后 2。早期的星形胶质细胞由脑室区的放射状胶质细胞产生,晚期则由室管膜下区的迁移祖细胞产生 2。在啮齿动物中,皮层星形胶质细胞主要由出生后前三周内局部分化的星形胶质细胞分裂产生 2。在人类中,星形胶质细胞的生成始于受孕后约24周,并持续到儿童早期 2。
调控机制
胶质发生在神经发生期间受到抑制。在神经发生结束时,促神经因子(如Ngn1)和抗星形胶质生成生长因子(如ERBB2)的下调促进了星形胶质细胞的产生 2。Notch、BMPs和IL-6家族细胞因子(LIF、cardiotriphin-1 (CT1) 和睫状神经营养因子 (CNTF))通过JAK-STAT3通路促进星形胶质细胞分化 2。表观遗传机制,如DNA甲基转移酶DNMT1介导的GFAP启动子CpG甲基化,也抑制星形胶质细胞生成 2。
在髓鞘化中的作用
星形胶质细胞在生理条件下积极促进髓鞘化,通过释放促髓鞘化因子、转运脂质和通过缝隙连接进行信号传导 34。
促髓鞘化因子: 星形胶质细胞是血小板衍生生长因子(PDGF)的主要生产者,PDGF通过其受体PDGFRα促进OPC增殖和迁移,并抑制OPC向成熟少突胶质细胞分化 34。星形胶质细胞还分泌脑源性神经营养因子(BDNF),在出生后早期发育中增强髓鞘化 34。睫状神经营养因子(CNTF)和白血病抑制因子(LIF)也具有强大的促髓鞘化作用 34。脂质转运: 髓鞘膜的形成需要少突胶质细胞合成大量脂质和蛋白质 41。研究发现,在正常大脑发育中,相当一部分整合到中枢神经系统髓鞘中的脂质是由星形胶质细胞提供的 41。星形胶质细胞衍生的胆固醇和脂肪酸对髓鞘形成至关重要 38。当少突胶质细胞或星形胶质细胞的脂质合成受损时,髓鞘形成会减少,但少突胶质细胞可以通过利用饮食中的脂质来弥补星形胶质细胞的缺陷 41。缝隙连接: 星形胶质细胞通过缝隙连接相互连接,并响应ATP提升细胞内钙离子浓度 25。它们也通过缝隙连接与少突胶质细胞进行信号传导,从而影响髓鞘化 36。
更广泛的作用
星形胶质细胞在塑造神经发育中发挥关键作用,包括促进神经元存活、迁移和轴突引导 2。它们最重要的功能之一是参与突触形成、成熟和修剪,以及神经回路的发育,控制兴奋性和抑制性突触发生 2。
疾病关联
星形胶质细胞生成缺陷与神经发育和精神疾病有关,这些疾病通常伴有突触发育和功能受损(例如,Rett综合征、脆性X综合征、唐氏综合征、自闭症、精神分裂症) 2。
星形胶质细胞在髓鞘化和神经回路精细化中扮演着积极且多方面的角色,远超简单的支持作用 25。它们通过分泌促髓鞘化因子、转运脂质以及通过缝隙连接进行信号传导,直接影响少突胶质细胞的分化和髓鞘的形成。这种多功能性挑战了神经功能以神经元为中心的传统观点,强调了神经胶质细胞在塑造大脑结构和功能中的根本重要性。例如,星形胶质细胞能够提供髓鞘形成所需的大量脂质,这表明它们是髓鞘生物合成的关键贡献者,而不仅仅是少突胶质细胞的辅助者。这种对胶质细胞主动作用的认识,为神经发育障碍和神经退行性疾病的治疗策略开辟了新途径,可能通过靶向胶质细胞功能来促进神经修复和功能恢复。
D. 小胶质细胞在神经系统发育中的作用
起源
小胶质细胞是中枢神经系统(CNS)的常驻巨噬细胞,起源于胚胎早期卵黄囊中的造血干细胞,并在神经前体细胞开始分化后不久定居于大脑 25。
作用
小胶质细胞在神经系统发育中发挥多方面作用,包括免疫监视、吞噬作用以及在神经炎症过程中的突出作用 25。它们不断地监测大脑和脊髓的所有区域,包括连接神经元的白质纤维束 31。
髓鞘化调控
小胶质细胞通过吞噬过多的少突胶质细胞和髓鞘膜,以及分泌营养因子、趋化因子和细胞因子来调节少突胶质细胞的分化和髓鞘化 31。
营养因子: 胰岛素样生长因子1(IGF-1)是小胶质细胞衍生的重要因子,支持少突胶质细胞健康,并显著减少培养中少突胶质细胞的细胞凋亡 31。小胶质细胞衍生的IGF-2也被证明可以防止肿瘤坏死因子-α(TNF-α)诱导的少突胶质细胞凋亡 31。细胞因子和趋化因子: 抑制小胶质细胞活化会损害少突胶质细胞的成熟,通过减少IL-1β和IL-6等小胶质细胞细胞因子的水平 31。早期出生后白质中的小胶质细胞,其转录特征与疾病状态下的小胶质细胞相似,具有促炎细胞因子和趋化因子(如Mif、Ccl3、Ccl4、CCl6、Ccl9)的基因转录本升高 31。其他细胞间信号分子: 跨谷氨酰胺酶-2(TG2)和小胶质细胞衍生的神经纤毛蛋白-1(NP1)也被发现支持OPC的增殖 31。
小胶质细胞能够感知神经元活动和微环境的分子特征,从而优化髓鞘化 31。
突触修剪
小胶质细胞还参与突触消除和回路精细化,有时通过补体系统进行 2。它们通过吞噬作用清除退化的神经元、髓鞘碎片和病理性tau蛋白聚集 31。
小胶质细胞在塑造神经回路中扮演着积极的角色,通过吞噬作用(清除多余的细胞、突触和髓鞘)和分泌调节分子来发挥作用 2。这表明小胶质细胞的功能远不止于免疫应答,它们直接参与了发育过程中的神经结构塑造。例如,小胶质细胞通过清除多余的少突胶质细胞和异常形成的髓鞘膜来优化髓鞘化,同时通过释放多种营养因子和细胞因子来支持少突胶质细胞的存活和分化。这种对小胶质细胞在正常发育中复杂作用的理解,为神经发育异常提供了新的视角,并可能为干预策略提供信息,以促进健康的白质发育和神经回路功能。
五、人类大脑和脊髓的区域发育
A. 大脑皮层发育
层状结构
大脑皮层,在哺乳动物中也被称为新皮层,在胚胎大脑最前部的背侧端脑中发育 2。该区域负责高级认知功能,并具有复杂的细胞结构,包括在该区域出生并投射的投射神经元,以及源自腹侧端脑区域的中间神经元 2。皮层投射神经元组织成六个层,形成称为皮层板的层状结构 2。这些层状神经元通过特定的分子标记组合和独特的轴突投射来区分 2。深层(V和VI)主要由投射到皮层下区域(如丘脑、脑干和脊髓)的皮层下行神经元组成 2。浅层(IV到II)由局部投射或投射到对侧半球的皮层内神经元组成 2。
神经元起源
所有这些兴奋性谷氨酸能神经元都源自发育中新皮层的生发区:脑室区(VZ),其衬里脑室;以及室管膜下区(SVZ),其从VZ发育而来并紧邻其基底表面 2。神经发生的第一阶段发生在VZ中,产生先驱神经元,包括定居于前板的Cajal-Retzius细胞 2。第二阶段,产生更多重要的神经元输出,主要发生在SVZ中,产生投射神经元 2。这些初级神经元将前板分裂为边缘带(第一层)和下板。下板是一个中间结构,神经元在迁移到其目标层时接收重要的信息线索 2。顺序产生的投射神经元以“由内向外”的方式迁移到皮层板,这意味着最年轻的上层神经元迁移到较早出生的深层神经元之上 2。这种迁移沿着放射状胶质细胞(RGCs)的基底突起进行,形成迁移神经元的放射状单元,这是大脑皮层径向扩展的主要过程 2。
脑回形成(Gyrification)
大脑皮层的体积在哺乳动物物种间差异显著 2。其进化上的增大与认知功能和智力的增强相关,并与脑回或脑沟的获得有关 2。这种折叠区分了灵长类动物(回脑)和啮齿动物(平滑脑)的皮层 2。皮层生长依赖于祖细胞的扩增,特别是在外室管膜下区(OSVZ),并根据物种的认知能力和感觉运动技能进行区域性变化 2。脑回形成的机制涉及细胞外基质成分和miRNA在控制OSVZ祖细胞扩增中的重要作用 2。TRNP1表达的变异与胎儿人脑室表面的区域性扩张相关 2。细胞骨架基因,特别是微管蛋白的突变,导致无脑回畸形,这说明了细胞迁移过程在脑回形成中的重要性 2。胚胎丘脑轴突提供重要线索,通过调节神经发生(刺激祖细胞增殖)和通过轴突张力产生力来影响皮层发育 2。这些力可能通过将皮层相互连接的区域拉到一起来影响脑回形成,从而可能优化皮层布线 2。丘脑传入轴突的早期局灶性消融可导致脑沟发育和方向的显著异常 2。
B. 小脑发育
整体结构与功能
小脑是中枢神经系统的重要组成部分,对熟练的运动表现至关重要,同时也参与认知和社会功能 43。它在内侧-外侧轴上分为四个主要区域:蚓部(和旁蚓部)、半球、绒球和旁绒球 44。
细胞起源
小脑的细胞起源复杂且精确。浦肯野细胞(Purkinje cells),作为GABA能(抑制性)神经元,起源于脑室区 43。而颗粒细胞(granule cells),作为谷氨酸能(兴奋性)神经元,以及小脑核神经元则来源于上菱唇 43。中间神经元和星形胶质细胞则由表达Nestin的祖细胞(NEPs)产生 43。
神经发生时间线
浦肯野细胞是小脑中最早生成的主要细胞类型之一,在小鼠胚胎第10天(人类受孕后约30天)开始生成 44。而颗粒细胞的神经发生高峰则发生在出生后,在人类中约为受孕后20-40周 44。
颗粒细胞迁移与分层
颗粒细胞前体细胞形成一个次级生发基质,称为外颗粒层(EGL),它覆盖在小脑原基的表面 43。在那里,前体细胞增殖数周,然后产生分化的颗粒细胞 44。颗粒细胞最终迁移到内颗粒层(IGL) 44。在此迁移期间,它们还会延伸平行纤维轴突,与浦肯野细胞形成突触连接 44。伯格曼胶质细胞(Bergmann glia),源自放射状胶质细胞,为颗粒细胞的放射状迁移提供支架 19。分子相互作用,如NRG-ErbB信号、FGF-FGFR1/2信号以及Mllt11和NMIIB等,在伯格曼胶质细胞的特性(如分化和成熟)中发挥作用,从而介导颗粒神经元的放射状迁移 15。
浦肯野细胞发育及其与颗粒细胞的相互作用
浦肯野细胞是小脑回路的核心,也是小脑皮层的唯一输出 44。它们被认为是小脑发育的“主控者”,在发育过程中引导传入纤维和中间神经元的布线 44。浦肯野细胞表达Sonic Hedgehog(SHH),这是一种对颗粒细胞前体扩增以及NEPs扩增和中间神经元及胶质细胞产生至关重要的有丝分裂原 43。
树突树形成: 在小脑皮层发育过程中,每个浦肯野细胞会生长出多个树突树,其中一个初级树突树被选择进一步发育,而其他树突树则完全退缩 47。这个选择过程由与颗粒细胞的物理和突触相互作用协调 47。颗粒细胞提供机械、分子和基于活动的线索,塑造浦肯野细胞的结构、连接和功能成熟 44。分子线索: 除了颗粒细胞与浦肯野细胞突触处的神经元通信外,其他颗粒细胞衍生的线索对于浦肯野细胞的最终成熟也是必需的 44。这些线索包括克隆细胞扩增产生的机械力以及颗粒细胞到浦肯野细胞的异源分子信号 44。攀援纤维输入对于取代竞争性传入和维持其树突区域至关重要 48。
小脑具有独特且漫长的出生后发育过程,特别是颗粒细胞的神经发生和迁移,以及浦肯野细胞作为“主控者”通过SHH信号传导及其与颗粒细胞的相互作用来协调这一复杂发育的关键作用 43。浦肯野细胞在胚胎期早期诞生,而颗粒细胞的大量增殖和迁移则主要发生在出生后。这种时间上的错位使得浦肯野细胞能够通过SHH等分子信号,在颗粒细胞群体扩张中发挥指导作用,并进一步通过与颗粒细胞的物理和突触互动,精细调整自身的树突树形态和回路连接。这种精密的协调对于小脑最终实现其在运动控制、学习和认知功能方面的多样化作用至关重要。对这些相互作用机制的理解,为小脑发育障碍的病理生理学提供了深入见解,并可能为未来靶向性干预策略提供基础。
C. 脑干发育
结构与功能
脑干是连接大脑和脊髓的神经组织束 49。它位于脊髓顶部,控制着呼吸、心跳、血压、意识和睡眠等维持生命的基本自主功能 13。脑干包含白质和灰质的集合,灰质由神经细胞体组成,形成许多重要的脑干核团 50。
区域划分
脑干由三个主要区域组成:中脑(mesencephalon)、脑桥(pons)和延髓(medulla oblongata) 49。
脑神经核团
12对脑神经中有10对起源于脑干 50。在胚胎发育过程中,脑神经核团在第四脑室底部的灰质中呈垂直排列,形成一系列称为核柱的结构 52。这些核柱根据其功能(运动或感觉)和相对于第四脑室底中线(正中沟)和界沟(sulcus limitans)的位置进行组织 52。例如,一般躯体传出(GSE)核柱最靠近正中沟,而特殊内脏传出(SVE)和一般内脏传出(GVE)核柱则更靠近界沟 52。随着发育的进行,这些核柱的某些部分会从其原始位置消失,或迁移到脑干深处,形成离散的核团 52。
主要传导束
脑干是许多重要神经传导束的通路。皮质脊髓束(运动)和脊髓丘脑束(感觉)都穿过脑干 13。许多运动神经束在延髓的锥体处交叉(锥体交叉),这意味着大脑的一侧控制身体的对侧 13。延髓也是控制胸腹部器官、肩部和头部运动、吞咽、唾液分泌、味觉、听觉和平衡的几对神经的起源 13。脑桥作为下脑干和中脑之间的“桥梁”,将神经冲动传递到小脑以协调复杂的肌肉运动 13。中脑主要作为感觉和运动神经冲动在脑桥、脊髓、丘脑和大脑皮层之间的中继中心 13。
特定核团
脑干还包含许多其他重要的核团:
红核: 位于中脑前内侧,参与运动功能,特别是上肢的精细运动,是下行红核脊髓束的起源 50。黑质: 位于中脑基底部,富含多巴胺能神经元,是基底神经节的一部分,在调节运动和奖励功能中发挥关键作用 50。帕金森病中多巴胺生成细胞的死亡导致运动功能障碍 50。橄榄体: 位于延髓,分为下橄榄核(协调脊髓和小脑之间的信息,调节精细运动控制和运动协调,可能参与运动学习)和上橄榄核(参与听觉神经系统,协助声音感知) 50。
D. 脊髓发育
结构与功能
脊髓是一个圆柱形、组织良好的结构,从颅底的枕骨大孔处作为延髓的延续开始 54。它位于椎管内,在成人中延伸至第一或第二腰椎水平,在儿童中则终止于第三腰椎上缘 54。脊髓分为31个节段,每个节段对应一对脊神经及其脊神经节 54。脊神经包含运动、感觉和自主神经纤维 54。
脊髓由白质和灰质组成 54。灰质是细胞体的集合,呈“H”形或“蝴蝶”形,位于脊髓中央 55。白质是轴突的集合,围绕灰质,并分为三个索 54。脊髓还被三层脑膜(硬脑膜、蛛网膜、软脑膜)包裹和保护 49。中央管贯穿灰质两半之间,与第四脑室相连,含有脑脊液 54。
生长差异
在胚胎发育的最初三个月,脊髓与椎管以大致相同的速度生长 55。然而,随着发育的继续,身体和椎柱的生长速度远快于脊髓本身 55。这种不均匀的生长导致成人脊髓终止于第一或第二腰椎水平,而神经根则向下延伸以从其适当的椎间孔穿出 55。
灰质组织
脊髓灰质的组织结构精细,不同区域具有特定功能:
背角: 存在于所有脊髓水平,由感觉核团组成,接收和处理传入的躯体感觉信息 55。中间柱和侧角: 包含支配内脏和盆腔器官的自主神经元 55。腹角: 包含支配骨骼肌的运动神经元 55。板层: 灰质根据Rexed板层进行组织。例如,板层III和IV对应于固有核,是脊髓丘脑束的突触部位 54。板层VI处理屈曲反射,并接收来自肌梭的信息,与板层VIII协同协调脊髓内的反射 54。板层VII包含Clarke背核,其产生同侧脊髓小脑束,将无意识本体感觉信息传递到小脑 54。
主要传导束与寻径
皮质脊髓束(CST): CST是提供随意运动功能的主要神经通路,起源于额顶叶皮层(包括初级运动皮层、次级运动区和躯体感觉皮层),然后通过内囊和大脑脚下行 56。其大部分纤维在延髓处交叉(锥体交叉)到对侧,然后继续下行到脊髓,支配远端肢体和肌肉群 56。在胚胎发育过程中,皮质脊髓束的轴突过度生长,并在发育过程中消除多余的轴突,从而进行精细化 56。这种精细化在出生后持续进行,并通过青春期受到雄激素水平上升的影响 56。轴突寻径由特定的分子信号事件精确引导,包括
Netrin-1、Plexin A、Semaphorins和Ephrins等引导分子,它们介导吸引和排斥作用,并指导中线交叉 17。Netrin-1的突变可能导致CST的异常交叉 57。CST结构的改变可能导致运动控制障碍 56。
脊髓丘脑束: 脊髓丘脑束传递疼痛、温度、粗略触觉和压力等感觉信息 50。侧脊髓丘脑束传递疼痛和温度,其纤维通过背角进入脊髓并早期交叉到对侧,位于侧索的深层 58。前脊髓丘脑束传递粗略触觉和压力,也进入后角并交叉,沿前脊髓丘脑束上行 58。Netrins在脊髓中线处引导这些轴突的交叉 17。脊髓小脑束: 传递无意识本体感觉信息,例如来自Clarke背核的纤维,其在同侧上行 54。
主要脊髓传导束(皮质脊髓束和脊髓丘脑束)的布线涉及到精密的引导机制,其中特定的轴突引导分子(如Netrins、Semaphorins、Ephrins)在介导吸引、排斥和中线交叉方面发挥着关键作用 17。Netrin-1作为一种分泌蛋白,在CNS中线处引导轴突交叉,例如在脊髓丘脑束中 17。Semaphorins和Ephrins则可能根据受体复合物、神经元亚型和发育时间,表现出吸引或排斥作用 18。对这些分子机制的理解揭示了神经回路形成的高度特异性。当这些引导机制发生中断时,例如Netrin-1突变导致皮质脊髓束异常交叉,便会导致严重的神经功能缺陷 57。这种认识不仅加深了对正常神经发育的理解,也为神经损伤后的再生策略提供了潜在靶点,通过调节这些引导分子来促进受损轴突的正确重连接。
六、环境和遗传对发育的影响
注:在关注了脑发育的时间线及分子机制后,我认为我们要重点关注大脑的可塑性,神经突触可塑性,神经连接的可塑性,从而来考虑一些精神情绪疾病的认知与行为治疗。
A. 关键期与大脑可塑性
大脑发育并非一个线性的、预设的过程,而是高度动态且受经验影响的。关键期是指婴儿期和儿童早期大脑对学习特定技能和响应环境特别开放的短暂而强烈的时期 59。在这些时期,大脑迅速建立支持未来学习、行为和情绪健康的神经连接 59。例如,如果婴儿在早期没有获得足够的口语输入,可能会影响大脑未来处理词语的能力;如果眼睛在早期没有获得正常的视觉输入,视觉系统的某些部分可能无法正常发育 59。同样,如果儿童在情绪低落时没有得到足够的抱起、安抚或安慰,大脑的应激反应系统可能会以不健康的方式发展 59。
积极的经验,如语言、社会互动和感官刺激,在这些关键期内塑造儿童的认知、情感和身体成长 59。与此相反,错过这些经验可能导致在生命后期更难解决的发育挑战 59。照护者与儿童之间响应性的“服务与回馈”互动,是影响儿童社会环境的关键因素,能够促进神经连接的形成 59。例如,当婴儿哭泣时,抱起并安抚他们可以建立信任并帮助调节大脑的应激反应系统;当婴儿发出咿呀学语的声音时,回应他们可以促进语言发展和社会互动 59。这些看似微小的行为,在关键期内对大脑区域的连接和生长产生巨大影响 59。
尽管错过关键期会使后期的学习变得更加困难,但大脑并非完全关闭了学习的大门。这种灵活性被称为大脑可塑性,它允许大脑响应新经验进行调整和重塑,尤其是在额外的支持或干预下 59。然而,关键期后的学习通常需要更多的努力,并且可能无法达到相同的技能水平 59。
B. 负面环境因素的影响
负面环境因素可能阻碍大脑的最佳发育 4。
毒性压力: 持续的毒性压力,例如忽视或虐待,可能导致儿童大脑应激反应系统的不健康发展 59。社会隔离: 动物研究表明,婴儿期的社会隔离会导致持久的情绪和行为困难 59。孕期母体因素: 孕期发生的各种母体因素可能导致神经发育障碍,包括低出生体重、早产以及胎儿暴露于酒精、药物或铅等污染物 61。孕期感染,如巨细胞病毒、风疹、疱疹、梅毒、弓形虫病和寨卡病毒感染,以及宫内感染,都可能损害胎儿发育中的大脑 62。暴露于甲基汞等毒素也是一个风险因素 62。新生儿疾病: 新生儿期的疾病,如细菌性脑膜炎、病毒性脑炎、严重或未经治疗的黄疸、脑出血以及脑部缺氧,都可能显著增加脑瘫的风险 62。出生后因素: 婴儿期和儿童早期的营养不良和环境剥夺(缺乏成长、发育和社会适应所需的身体、情感和认知支持)可能是全球范围内智力障碍最常见的原因 64。
C. 遗传因素与神经发育障碍
神经发育障碍是一组影响大脑和神经系统发育方式的疾病 61。它们通常在成年期之前开始,并可能持续到成年期,常伴有社交障碍和沟通困难 61。这些疾病的精确病因尚不完全清楚,但研究表明遗传和环境因素的结合可能发挥作用 61。
神经管缺陷(NTDs)
定义: 神经管缺陷是脊髓或大脑的畸形,由胚胎神经系统(神经管)在出生前未能完全闭合引起 1。常见的类型包括脊柱裂(如脊髓脊膜膨出)和无脑畸形 65。时间: 神经管在胚胎发育的早期形成,通常在受孕后一个月内,有时甚至在母亲意识到怀孕之前 65。病因: NTDs被认为是复杂疾病,由多种基因和环境因素共同作用引起 65。已知环境因素包括叶酸缺乏(通过补充叶酸可预防50-70%的NTDs)、母体胰岛素依赖性糖尿病和母体使用某些抗惊厥药物 65。遗传因素也发挥作用,同卵双胞胎比异卵双胞胎更容易同时患有NTDs,且家族中已有患儿的夫妇再次生育患儿的风险增加 65。NTDs也是某些已知遗传综合征的特征,如13三体综合征、18三体综合征和Meckel-Gruber综合征 65。诊断与治疗: 大多数NTDs可在孕期通过高分辨率超声或羊膜穿刺术检测 65。出生后,通常需要早期手术修复以防止感染和神经功能缺损 67。
自闭症谱系障碍(ASD)
定义: ASD是一种复杂的神经发育障碍,涉及思维、情感、语言和身体能力(如运动和协调)的变化 61。其特征是社交互动和沟通困难,以及受限或重复的行为 61。ASD在胎儿发育和儿童早期形成,具有早期发病和强烈的遗传成分 68。神经生物学基础: ASD由特定基因的表达改变引起,并涉及转录因子和染色质重塑过程在脑神经元特定复合体内的特异性相互作用 68。突触,特别是树突的突触后部分,是ASD的早期和关键靶点 68。ASD在大脑灰质中分布广泛,包括中脑、脑桥、双侧海马、左侧海马旁回以及各种颞叶和枕叶脑回 68。基因组研究发现ASD与其他疾病(如癫痫和精神分裂症)存在遗传相似性,这强化了ASD具有强烈神经发育成分的观点 69。研究还强调了发育中突触功能改变和GABA能神经传递在ASD中的重要性 69。叶酸受体α(FRα)自身抗体在ASD中普遍存在,它们通过结合FRα来破坏叶酸跨血脑屏障的转运,可能作为ASD亚组的生物标志物 69。
注意缺陷/多动障碍(ADHD)
定义: ADHD是一种常见的、长期的、可治疗的儿童精神疾病,其特征是与发育水平不符的注意力不集中、运动不安和冲动模式 61。约70%的患儿在青少年期仍患有此病,约40-60%的患儿在成年期仍受影响 70。神经生物学基础: ADHD并非单一的病理生理实体,其病因复杂,涉及多种遗传和环境风险因素的协同作用 66。结构影像学研究表明,ADHD患儿的大脑比未受影响的对照组小,特别是前额叶皮层、基底神经节和小脑受到不同程度的影响,且关键脑区白质束连接减少 66。神经科学家普遍认为ADHD患者主要存在神经递质多巴胺利用不当的问题,其次是去甲肾上腺素 70。多巴胺显著影响运动调节、认知、情绪和注意力等多种行为 70。神经科学研究描述了三种注意力网络:腹侧、背侧和默认执行控制,任何网络的失调都可能导致ADHD的临床表现 70。运动困难,如感觉运动协调问题、笨拙和运动里程碑延迟,在ADHD人群中也很常见,可能与小脑和基底神经节的结构或功能异常有关 66。
智力障碍(ID)
定义: 智力障碍(原称精神发育迟滞)是一种神经发育障碍,其特征是智力功能显著低于平均水平(通常智商低于70-75),并伴有适应功能(如沟通、自我指导、社交技能、自我护理、社区资源利用和维护个人安全)的限制 61。这些缺陷通常在儿童早期(通常在18岁之前)开始显现 64。病因: 智力是遗传和环境共同决定的 64。智力障碍的病因多种多样,可能与遗传综合征(如唐氏综合征、脆性X综合征、染色体异常)有关 64。产前因素包括母体药物和毒素暴露(如胎儿酒精综合征、抗惊厥药物、铅、甲基汞) 64。出生后因素如婴儿期和儿童早期的营养不良和环境剥夺可能是全球范围内最常见的原因 64。共病: 智力障碍个体常伴有其他神经发育、精神和身体疾病,包括自闭症谱系障碍、脑瘫、癫痫、注意缺陷/多动障碍和抑郁焦虑症 71。
学习障碍
定义: 学习障碍(或特异性学习障碍)是一种神经发育问题,指具有正常智力潜能(即没有智力障碍)的儿童在学习特定学术技能(阅读、书写、数学)方面遇到异常困难 61。这些困难可能影响语言、沟通、社交情感和行为等多个方面 72。病因: 学习障碍的病因尚不完全清楚,但通常具有家族遗传性,环境因素也可能发挥作用 72。类型: 主要类型包括阅读障碍(dyslexia)、书写障碍(dysgraphia)和计算障碍(dyscalculia) 73。
脑瘫(CP)
定义: 脑瘫是一组影响运动和姿势的疾病,由发育中的大脑受损引起,最常见于出生前 62。症状通常在婴儿期或学龄前出现,严重程度差异很大 62。病因: 脑瘫的发生是由于大脑控制肌肉运动的区域受损或未正常发育 63。这种损伤可能发生在出生前(约80%)、出生时或出生后早期(约10%) 62。多种因素可能导致大脑发育改变,包括基因改变、影响胎儿的母体感染、中风(中断发育中大脑的血液供应)、胎儿期或新生儿期脑出血、导致大脑或周围肿胀的婴儿感染、婴儿创伤性头部损伤以及分娩过程中脑部缺氧(尽管这种情况比以前认为的少见) 62。早产和低出生体重是重要的风险因素 63。症状: 脑瘫患儿可能表现出运动技能里程碑延迟(如坐立或爬行)、反射亢进、学习障碍、智力障碍、生长迟缓、癫痫发作、听力或视力问题、感觉障碍、膀胱和肠道问题以及心理健康问题 62。导致脑瘫的脑部状况不会随时间恶化,但某些症状可能随着年龄增长变得更明显或不明显 62。
神经发育障碍的发生,是遗传倾向与环境因素复杂相互作用的结果 2。特定的基因突变(例如,影响神经发生或迁移的基因)或环境损害(例如,母体感染、毒素暴露、营养缺乏)都可能破坏神经发育过程中精确的细胞和分子事件。这种破坏并非导致单一的疾病,而是可能产生一系列多样化的临床表型,从轻微的学习困难到严重的智力障碍或自闭症谱系障碍。这种“责任谱系”的概念表明,神经发育障碍的病因是多因素的,并且个体对这些因素的脆弱性存在差异。因此,理解和干预这些疾病需要多方面的综合方法,不仅要考虑遗传易感性,还要关注生命早期和关键发育窗口期的环境暴露和支持。
七、结论
人类神经系统的发育是一个极其复杂、漫长且高度协调的生物学过程。它从受精后神经板的诱导和神经管的形成开始,通过神经发生、神经元迁移、分化、突触发生、髓鞘化以及细胞死亡与修剪等一系列精密的细胞和分子事件,最终构建出具有高级认知和运动功能的复杂网络。这一过程的独特之处在于其持续时间之长,从胚胎期一直延伸到成年早期,这意味着神经系统在生命的大部分时间里都处于持续重塑和对内外环境敏感的状态。
报告中对神经诱导中BMP拮抗剂作用的阐述,揭示了神经命运的“默认”机制,强调了精确分子反向调节的重要性。对神经发生中核间运动和细胞分裂物理机制的分析,突显了细胞力学在塑造大脑结构中的关键作用。神经元迁移过程中引导线索的上下文依赖性和多功能性,展示了神经回路布线的惊人精度与潜在脆弱性。突触发生的早期启动,表明功能性连接在出生前已奠定基础,为后期精细化提供了起点。
此外,胶质细胞,特别是星形胶质细胞和小胶质细胞,被证明并非仅仅是神经元的被动支持者。星形胶质细胞通过分泌促髓鞘化因子、转运脂质和缝隙连接信号,积极参与髓鞘形成和回路精细化。小胶质细胞则通过吞噬作用清除多余细胞和突触,并分泌调节分子,直接参与神经回路的塑造。这些发现挑战了传统上以神经元为中心的观点,揭示了胶质细胞在神经发育和功能中的核心地位。
区域性发育的差异,如大脑皮层的分层和脑回形成、小脑独特的出生后发育模式以及脑干和脊髓中特定核团和传导束的精确布线,都依赖于复杂的细胞间相互作用和分子信号。任何这些过程中的中断,无论是遗传突变还是环境暴露,都可能导致一系列神经发育障碍,包括神经管缺陷、自闭症谱系障碍、注意缺陷/多动障碍、智力障碍、学习障碍和脑瘫。这些疾病的病因往往是多因素的,涉及遗传易感性和环境影响的复杂交织。
综上所述,人类神经系统的发育是一个高度动态、脆弱且适应性强的过程,其复杂性要求我们从多层次、多维度进行深入研究。对这些基本机制的持续探索,不仅能增进我们对健康大脑如何形成的理解,更能为预防、诊断和治疗神经发育障碍提供新的靶点和策略,最终改善受影响个体的生活质量。